


正常な胎盤及び胚の発育に必須の卵子X 染色体の活動を維持する仕組みを解明
不妊・不育症、X染色体に関連する難治疾患の発症機序解明の手掛かりとして期待
国立成育医療研究センター研究所の福田篤研究員と阿久津英憲生殖医療研究部長のグループは、受精後に卵子X染色体が優先的に働く仕組みを世界で初めて明らかにしました。
プレスリリースのポイント
- 受精後、卵子X染色体(Xm)全体の活性調節を担う「しるし」として、卵子X染色体上に位置しタンパク質をコードしない遺伝子であるXist発現を調節する、ヒストンの化学的修飾の存在意義を発見しました。
- 染色体不活性化の破綻は、着床後の正常な発生を阻害することから、原因不明の反復性流産との関連も示唆されており、こうした不妊・不育症に関わる疾患ではゲノムレベルでの異常が認められない症例も多く、今回の発見は後発性の原因不明な不妊・不育症を克服する突破口となり得ます。
- 卵巣腫瘍や乳癌などの女性腫瘍の発症機序解明、そして安全で効果的な再生医療のためのES細胞やiPS細胞の品質にも関わる重要な成果でもあり、再生医療の発展に大いに貢献するものであると考えます。
背景
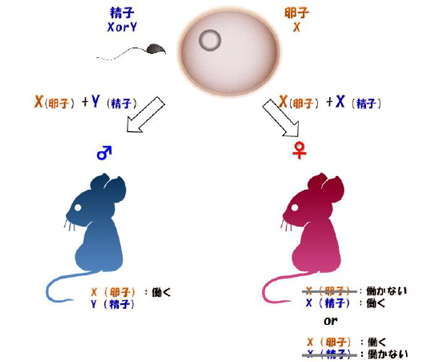
哺乳類の雌雄間では、性染色体であるX染色体の数が異なる。オス(1本のX染色体と1本のY染色体)では、常時X染色体が活性化しています。一方、メス(2本のX染色体)では、どちらか一方のX染色体が抑制されています(図1)。
|
図1:X 染色体の雌雄間における遺伝子発現量補正
 |
遺伝子の発現量が同じであることが生存には必須であり、X染色体に関しては、遺伝子発現量の補正機構が種を超えて存在しています(補足説明①)。この機構はX染色体不活性化とも言われ、中心的役割を担う分子としてXist(イグジスト)が知られています(補足説明②)。
哺乳類では、この現象が受精直後からみられ、正常な胎盤及び胚の発育には必須です。哺乳類のモデル動物では、受精後卵子由来のX染色体が優先的に働く仕組みが備わっています。しかし、正常な発生に極めて重要なその仕組みは明らかにされていませんでした。
研究手法と成果
この度、遺伝子発現量の補正を担う卵子X染色体に特異的に存在する「しるし」を同定しました。この「しるし」はDNAに結合する核(ヒストン)タンパク質の化学的修飾です(図2)。
|
図2:卵子X 染色体特異的な「しるし」
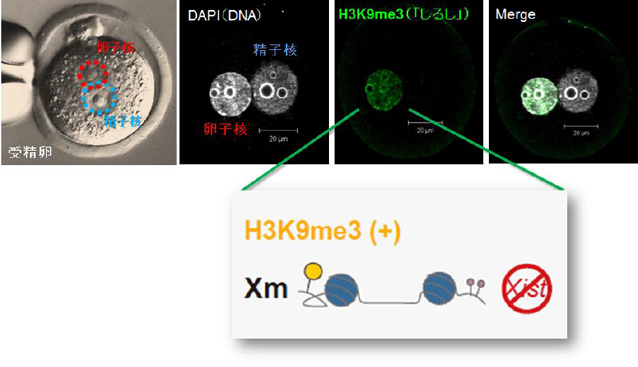 |
卵子にはXist 発現を調節するヒストンの化学的修飾(「しるし」)が付いている。この「しるし」は、ヒストン3(H3)の9 番目のリジン(K)(アミノ酸)にメチル基(me)が3 つ付加されていることである(H3K9me3)。これによりXist の発現が卵子側で抑制され卵子X 染色体(Xm)全体が活性化される。
哺乳類のX染色体は異なる種でほとんど同じ長さを持ち、含まれている遺伝子も共通なものが多い。例として、グルコース6リン酸脱水素酵素(G6PD:グルコース-6-リン酸脱水素酵素欠損症)やジストロフィン(dystrophin:筋ジストロフィー症)などがあり、様々な疾患と関連が深いとされています。
X染色体不活性化の破綻は、着床後の正常な発生を阻害することから、原因不明の反復性流産との関連も示唆されます。さらに、早発卵巣機能不全(Premature Ovarian Failure; POF)でもX染色体不活性化の乱れとも関連性が指摘されてきました。
これら疾患では、ゲノムレベルでの異常が認められない症例も多く、本研究成果でゲノムの外につく「しるし」を同定したことで後発性の原因不明不妊・不育症を克服する突破口となり得ます。
今後の展望・コメント
哺乳類の発生では、単為発生を妨げる防御機構が備わっていますが、ヒトでは、その防御機構の破綻から一部が卵巣内で腫瘍化することが知られています。また、乳癌などではX染色体不活性化と腫瘍発症との関連性が指摘され世界的に研究が活発化しています。したがって、本研究成果が、卵巣腫瘍や乳癌などの女性腫瘍の発症機序解明に繋がると考えられます。
また、ヒトiPS及びES細胞(46,XX染色体型)では、長期培養によりX染色体不活性化機構が破綻するとの報告があります(参考文献1)。今回の発見は、個体発生の源となる初期胚のX染色体不活性化制御機構を明らかにするものであり、多能性幹細胞での制御機構にも役立つことでしょう。
これにより、今回の研究成果は安全で効果的な再生医療のためのES細胞やiPS細胞の品質に関わる重要な成果でもあり、再生医療の発展に大いに貢献するものであると考えます。
発表論文情報
著者:Atsushi Fukuda, Junko Tomikawa, Takumi Miura, Kenichiro Hata, Kazuhiko Nakabayashi, Kevin Eggan, Hidenori Akutsu, Akihiro Umezawa.
題名:The Role of Maternal-Specific H3K9me3 Modification in Establishing Imprinted X-Chromosome Inactivation and Embryogenesis in Mice
掲載誌:Nature Communications, 2014, November 14
補足説明
- X染色体の遺伝子発現補正機構
哺乳類の雄では性染色体としてXとY染色体を1本ずつもち、雌はX染色体を2本もつ。雌の2つのX染色体のうちの一方が「X染色体不活性化(補足説明2)」という仕組みによって、働かない状態になっている。ショウジョウバエでは、雄(XY)のX染色体の遺伝子発現が雌(XX)のX染色体より2倍に高められている。一方、線虫では、雌雄同体(XX)の遺伝子発現量が、雄(XO)のX染色体の半分に抑えられている。
いずれの生物においても、このX染色体遺伝子発現機構が破綻すると致死となることから、X染色体の遺伝子発現補正が生き物の正常な発生にとって極めて重要である(参考文献2)。 - X染色体不活性化
X染色体上に位置するタンパク質をコードしない遺伝子であるXistが発現することで、そのX染色体全体の遺伝子発現が抑制される。雌の2本のX染色体の一方が不活性化される現象は、1961年にイギリスのメアリー・ライオン博士によって初めて報告されている。Xistは受精後から発現が開始されるが、その制御機構は長らく不明であった。
参考文献
(1) Mekhoubad S, Bock C, de Boer AS, Kiskinis E, Meissner A,EgganK. "Erosion of dosage compensation impacts human iPSC disease modeling."Cell Stem Cell. 2012; 10(5): 595-609.
(2) 佐渡敬 「X染色体不活性化のメカニズムを解く」 ,総研大ジャーナル9月号 ,2006; 国立大学法人 総合研究大学院大学
- 本件に関する取材連絡先
-
国立成育医療研究センター 企画戦略局 広報企画室
03-3416-0181(代表)
koho@ncchd.go.jp
月~金曜日(祝祭日を除く)9時〜17時
※医療関係者・報道関係者以外のお問い合わせは、受け付けておりません。